陈凯歌 男同 雅砻江冬麻豆(Salweenia bouffordiana)种群结构与动态特征

种群应系指一个地区内同属于一个种的个体组成的群体[1], 种群结构是种群最基本的特征, 是种群生态学磋议的中枢问题之一[2-3]。极小种群是指地舆漫衍短促的种群, 长久受到外界身分的侵犯和压力陈凯歌 男同, 其数目小于耀眼逝世所需的最低适度[4], 极小种群具有剩余种群少、栖息地受限、东谈主为侵犯严重和绝种风险极高的特征[5]。当今, 我国仅有120个极小种群被柔柔[4-5], 浩大处于濒危状态的物种并未得回保护地位, 而其算作生物各种性的首要组成, 一朝逝世将给东谈主类带来不行斟酌的赔本。面前, 物种逝世已是大家最严重的生态问题之一, 告成挟制着东谈主类社会的可捏续发展[6], 而保护濒危物种已成为一个紧迫的保护生物学课题[7]。中国算作宇宙上最大的生物各种性国度之一, 保护中国最受挟制的野生植物物种比以往任何时候皆愈加遑急。
雅砻江冬麻豆(Salweenia bouffordiana)是豆科冬麻豆属(Salweenia)植物新种[8], 为我国特有种, 主要漫衍于四川省新龙县境内的雅砻江河谷干燥灌丛和砾石生境中[8]。由于本种的漫衍界限极其轻飘, 种群数目疏淡, 把柄宇宙自然保护定约条例[9], 有学者们将其保护品级定为相当濒危(Critically Endangered, CR)[8], 《中国生物各种性红色名录——高档植物卷》将其定为濒危(Endangered, EN)物种[10]。由于短缺对该物种的种群数目、种群结构等方面的磋议, 不利于后续对于该物种的相关保护计谋的制定和行动的开展, 需要对其现有种群数目、漫衍界限等进行访谒与磋议。
1 材料与要津 1.1 磋议区概况雅砻江冬麻豆为常绿丛生灌木, 高约0.5—2 m, 茎耸峙, 被绒毛, 与冬麻豆(S. wardii)的辞别主要在于本种的小叶更大、瓣膜呈浪潮状, 主要漫衍于四川省新龙县境内的雅砻江河谷。本磋议的磋议区位于四川省甘孜藏族自治州新龙县, 地舆位置介于东经99°37′—100°54′E、30°23′—31°32′N之间, 海拔2760—3232 m, 地处雅砻江中游峻岭峡谷地带。该区属青藏高原型季惬心象, 具有专有的大陆性高原季惬心象的特质。把柄新龙县现象站多年统计尊府可知, 全县平均气温为7.5℃, 顶点最高温32.7℃, 顶点最低温-19.2℃, 1月均温-2.3℃, 7月均温15.1℃, ≥10℃行径积温1887℃, 相对湿度在40%独揽, 全年无霜期114 d。磋议区水量漫衍不均匀, 降雨主要聚会于每年6月至9月。植被群落主要有河谷灌丛、亚峻岭暗针叶林、峻岭松林、峻岭柏林、栎类林、桦木林、亚峻岭草甸、峻岭草甸等类型。
1.2 访谒要津由于雅砻江冬麻豆主要滋长在雅砻江河谷地带, 故沿河岸1 km界限偶然布设访谒样线8条, 沿每条样线记载发现的雅砻江冬麻豆、侵犯信息和生境信息。在有雅砻江冬麻豆种群漫衍的地段成立5 m×5 m样方作念成例的种群学访谒, 合计访谒样方55个。样方访谒时记载样方的经纬度、海拔、坡向、坡度等因子, 乔木层、灌木层和草本层的植物种类、盖度和高度等植被状态, 并对雅砻江冬麻豆的种群及滋长状态进行成例种群学访谒, 记载样方内雅砻江冬麻豆的幼苗数目、盖度、高度、冠幅和地径等参数。
1.3 斟酌要津 1.3.1 径级离别圭表对于木本植物尤其濒危物种而言, 大多数学者以为在数目较少, 较难获取其年事的情况下, 可用径级结构代替其年事结构[11]。本磋议遴荐生物方向特征显赫的地径结构来代替年事结构, 为分析其种群结构的缜密动态, 经受李清河等对零散疏淡灌木半日花种群的径级离别要津[12]进行地径离别, 即经受较小极差, 每1 mm离别1个径级。把柄地径(D)的漫衍状态, 将地径离别为25个品级:Ⅰ级(3 mm≤ D < 4 mm)、Ⅱ级(4 mm≤ D < 5 mm)、Ⅲ级(5 mm≤ D < 6 mm)、……、XXIV级(26 mm≤ D < 27 mm)和XXV级(D ≥27 mm)。以上述离别为圭表, 统计每个龄级雅砻江冬麻豆个体数, 并以此进行种群动态量化分析和设置种群静态人命表。
1.3.2 种群动态量化分析要津种群动态量化经受陈晓德[13]的量化分析要津定量描画雅砻江冬麻豆种群动态, 以Leak[14]的离别表面算作详情种群结构类型的依据, 公式如下:
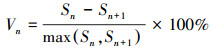 (1)
(1)
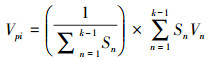 (2)
(2)
式中, Vn为种群从n 到n+1级的个体数目变化;式(2)中Vpi为悉数这个词种群结构的数目变化动态指数(忽略外部侵犯);Sn、Sn+1分别为第n和n+1级种群个体数;k为种群大小级数目。而当研究将来的外部侵犯时, Vpi还与年事级数目(k)以及各龄级内的个体数(Sn)相关, 可进一步将式(2)修正为:
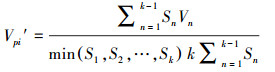 (3)
(3)
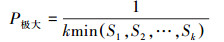 (4)
(4)
式中, Vn、Vpi、Vpi′取正、负、零值期间别反应种群个体数目的增长、阑珊和踏实的结构动态关系。其中, Vpi值越大, 诠释种群的增长趋势越大, 而Vpi′值越大, 诠释种群抗侵犯才智越大, 种群踏实性越好。仅当P(种群对外界侵犯所承担的风险概率)取值为最大时才会对种群动态Vpi′组成最大的影响。
1.3.3 种群静态人命表和活命分析要津静态人命表又称特定期间人命表, 多用于对龟龄命的木本植物种群的统计磋议, 主要包括以下参数:Ax是在x龄级内现有个体数;lx是在x龄级运转时圭表化存活个体数;dx是从x到x+1龄级阻隔期内圭表化死一火数;qx是从x到x+1龄级阻隔期间死一火率;Lx是从x到x+1龄级阻隔期间还存活的个体数;Tx是从x龄级到卓绝x龄级的个体总和;ex是干涉x龄级个体的人命生机或平均生机寿命;Kx为隐没率(赔本度)。它们之间的斟酌关系如下:
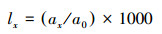 (5)
(5)
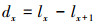 (6)
(6)
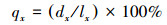 (7)
(7)
 (8)
(8)
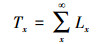 (9)
(9)
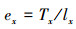 (10)
(10)
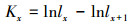 (11)
(11)
由于静态人命表是用归拢时期集聚的种群悉数个体编制而成, 雅砻江冬麻豆种群为自然种群, 分析中专揽不同龄级结构在空间上的互异来磋议特定期间段的种群动态, 不免存在系统抽样罪恶产生与数学假定时期不符的征象, 但仍能提供有用的生态学记载[15]。对于这种情况, 在编制静态人命表频繁经受匀滑时期对统计数据进行处理, 本磋议经受方程拟合要津进行匀滑处理[15-17]。由于第Ⅵ龄级前的植株数存在显明颠倒, 是以拟合经受第Ⅵ龄级特等之后的数据, 以年事级为自变量, 以龄级对应的现有个体数Ax为因变量, 经R3.3.3软件拟合得到方程:y=1473.506x-2.1263 (R2=0.7517, P < 0.001)。经此拟合方程匀滑修正后得到修正后的ax, 然后据此编制出雅砻江冬麻豆种群的静态人命表。
为了测验种群存活状态是安妥Deevey-Ⅱ型也曾Deevey-Ⅲ型弧线, 本文经受Hett和Loucks提倡的数学模子对雅砻江冬麻豆种群存活弧线进行测验, 即用指数方程Nx=N0e-bx和幂函数方程Nx=N0x-b分别描画Deevey-Ⅱ和Deevey-Ⅲ型存活弧线[18]。式中Nx、N0分别代表x龄级内存活数和种群形成初期的个体数, 即Nx、N0为经匀滑后的个体存活数陈凯歌 男同, b为死一火率。
为了更好地分析雅砻江冬麻豆种群动态, 报酬其活命限定, 经受种群活命率函数(S(i))、累计死一火率函数(F(i))、死一火率密度函数(f(ti))和危急率函数(λ(ti))进行分析[19-20]。斟酌公式如下:
bt工厂最新地址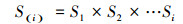 (12)
(12)
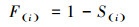 (13)
(13)
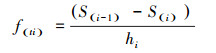 (14)
(14)
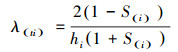 (15)
(15)
式中, Si为存活率, 其斟酌公式为:Si=lx+1/lx, hi为龄级宽度。依据这4个活命函数的估算值, 绘画活命率弧线、累计死一火率弧线、死一火密度弧线与危急率弧线。
1.3.4 种群数目动态的期间序列忖度要津本磋议经受期间序列分析中的一次转移平均法[21]对雅砻江冬麻豆种群的年事结构进行忖度。
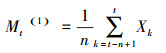 (16)
(16)
式中, n暗意需要忖度的期间(本磋议为龄级期间), t为龄级, Xk为k龄级内的个体数目, Mt(1)暗意经过将来n个龄级期间后t龄级的种群大小。本文对将来经过2、4、9、6、8、12、20和24龄级期间后的种群各龄级的个体数目进行忖度。
2 服从与分析 2.1 雅砻江冬麻豆种群漫衍近况和结构特征雅砻江冬麻豆种群年事结构呈纺锤形(图 1)。种群植株个体主要聚会在第Ⅲ—XIV龄级, 即地径5—17 mm之间, 占总植株数的80.71%。其中, 地径8—15 mm的植株数占总植株数的56.85%, 诠释第Ⅵ—Ⅻ龄级的植株数目充足, 而地径3—8 mm和>17 mm之间的植株数分别占总植株数的18.27%和14.72%, 诠释雅砻江冬麻豆种群短缺低龄级和乐龄级植株, 且第Ⅰ—Ⅲ龄级的植株个体数占比之和不及9%, 第XXIII—XXV级的植株个体数占比之和不及3%, 标明种群有一定的更新才智, 但更新潜力不及, 甚为短缺龄级两头处的植株, 种群呈现出阑珊状态。
把柄访谒样方幼苗数统计袒露, 约53.57%的样方中有雅砻江冬麻豆幼苗, 其中以1—5株幼苗的居多, 占总样方数的39.29%(图 2), 标明磋议区内幼苗虽有漫衍, 但其个体数目仍相当短缺, 种群更新才智低下。雅砻江冬麻豆种群合座呈现出中龄数目丰富, 低龄数目短缺, 乐龄数目严重不及的纺锤形结构, 种群更新不良, 种群年事结构为阑珊型。
由表 1可知, 雅砻江冬麻豆种群V6、V17、V19、V23均就是0, 诠释第Ⅵ—Ⅶ、XVII—XVIII、XIX—XX、XXIII—XXIV龄级间的个体数目莫得变化, V4、V8、V10、V12、V14、V15、V16、V18、V20均大于0, 诠释第Ⅳ—Ⅴ、Ⅷ—Ⅸ、Ⅹ—Ⅺ、Ⅻ—XIII、XIV—XVII、XVIII—XIX、XX—XXI龄级间的个体数目呈现增长的结构动态关系, 而剩余的龄级间均呈现出个体数目减少的结构动态关系, 标明种群总体呈现出“阑珊—增长—阑珊—增长”的来往波动性结构动态关系。进一步分析其数目动态变化指数Vpi(不研究外部侵犯)和Vpi′(研究偶然侵犯)的服从袒露, Vpi=0.0799, Vpi′=0.0032, Vpi > Vpi′, 自然种群总体动态变化指数Vpi大于0, 但仅为0.0799, 且Vpi′值仅为0.0032, 均趋向于0, 标明种群波动大, 种群踏实性差, 抗侵犯才智低。偶然侵犯风险极大值P极大=0.04, 即种群结构对偶然侵犯的敏锐性指数为0.04, 诠释雅砻江冬麻豆种群对外界偶然侵犯具有较高的敏锐性, 标明其对环境的顺应力低, 何况由于种群结构低龄个体更新不良, 因此雅砻江冬麻豆种群应属于阑珊型种群。
2.2 雅砻江冬麻豆种群静态人命表由雅砻江冬麻豆种群静态人命表可知, 跟着年事的增大, 雅砻江冬麻豆种群存活数(lx)缓缓减小(表 2)。植株个体生机寿命(ex)跟着龄级的加多呈现出先增后减的状态, 其反应了植株个体的人命生机寿命, 其最小生机寿命在第XXV龄级处(ex =1), 最大生机寿命在第XIV龄级处(ex =7.667), 诠释雅砻江冬麻豆种群在较乐龄级阶段仍然具有较高的人命生机。种群死一火率(qx)与隐没率(Kx)反应了种群数目跟着龄级加多的动态变化, 雅砻江冬麻豆种群死一火率与隐没率在第Ⅻ龄级前均随龄级加多而呈加多的状态, 但在第Ⅻ龄级特等后出现剧烈波动, 标明种群处于极不踏实状态。因此, 雅砻江冬麻豆种群合座呈现出极不踏实的状态, 种群发展状态差。
经R3.3.3软件拟合, 雅砻江冬麻豆种群存活弧线的拟合方程为:
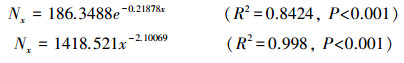
模子测验服从标明, 两种模子的拟合服从在雅砻江冬麻豆种群中均达到了极显赫水平, 由于幂函数模子的R2值比指数函数模子大, 故以为雅砻江冬麻豆种群存活弧线更趋近于Deevey-Ⅲ型。依据Deevey的离别[22]:Ⅰ型呈凸弧线, 多数个体皆能活到该物种的生理年事, 但当达到一定生理年事时则广泛死一火;Ⅱ型呈直线, 诠释各年事的死一火率相当;Ⅲ型是凹弧线, 早期死一火率极高。因此雅砻江冬麻豆种群具有早期死一火率极高的特质。
以龄级为横坐标, 以存活数的对数为纵坐标, 把柄雅砻江冬麻豆种群静态人命表绘画存活弧线(图 3)。由图 3可知, 雅砻江冬麻豆种群存活弧线在第Ⅰ到第Ⅲ龄级险些成直线下落, 干涉第Ⅳ龄级后, 缓缓趋缓, 且变化率在第XV、XVI、XIX龄级处和第XXII龄级特等之后均为0, 同期自第XXI龄级之后存活数的对数值也均为0, 标明雅砻江冬麻豆种群低龄级死一火率高, 中龄级植株存活率相对较高, 当种群干涉第XV、XVI、XIX龄级登科XXI龄级之后大部分植株会较快死一火。在存活弧线方面, 雅砻江冬麻豆种群与濒危物种鹅掌楸、矮牡丹、裂叶沙参、木根麦冬、长喙毛茛泽泻种群的磋议服从一致[23], 均为Deevey-Ⅲ型, 高死一火率聚会在低龄阶段, 具有生殖才智的成年个体少。
2.3 雅砻江冬麻豆种群活命由图 4可知, 雅砻江冬麻豆种群的活命率随龄级增大发扬出先急剧减小此后趋缓的状态, 累计死一火率呈现出先快速递加此后趋缓的态势, 二者互补, 但种群活命率远小于累计死一火率, 标明种群处于阑珊状态。活命率弧线和累计死一火率弧线在前3个龄级呈现出急剧升降变化, 在第Ⅴ龄级后缓缓趋缓, 这可能和种群在低龄级的高死一火率联系, 广泛低龄级个体死一火, 种群呈现阑珊特征。在第Ⅵ龄级处, 种群活命率为0.016, 累计死一火率为0.984, 种群已干涉阑珊期, 标明雅砻江冬麻豆种群在活命早期已处于阑珊状态。
种群死一火密度弧线和危急概率弧线与其活命率和累计死一火率变化趋势一样, 均在前3个龄级急巨变化, 但种群危急概率弧线一直显赫大于死一火密度弧线, 在干涉第Ⅳ龄级后均变化趋缓, 其中危急概率弧线的合座变化更显赫, 死一火密度弧线在第XXV龄级达到最小值(0), 危急概率弧线在第XXV龄级达到最大值(2)。由4个活命函数弧线标明, 雅砻江冬麻豆种群具有前期薄弱, 中期、后期阑珊显赫的特质, 种群活命力较低, 处于阑珊状态。
2.4 雅砻江冬麻豆种群数目的期间序列雅砻江冬麻豆种群数目动态变化的期间序列忖度是以其种群各龄级株数为原始数据, 按照一次转移平均法忖度出各龄级在将来2、4、9、6、8、12、20和24个龄级期间后的个体数。从表 3分析可知, 资格2、4、6个龄级期间, 种群从第Ⅻ龄级运转之后均有不同进度的加多, 经过8和12个龄级期间, 从第Ⅻ龄级运转减少, 此后趋于踏实, 诠释一朝过了一定的生理年事, 雅砻江冬麻豆种群就运转缓缓走向阑珊。种群前期, 在第Ⅵ龄级经过2、4和6个龄级期间后个体数分别下落了21.43%、28.57%和42.86%;种群前中期, 在第Ⅺ龄级经过2、4、6和8个龄级期间后个体数下落了16.67%、25.00%、37.50%和41.67%;而种群中期, 在第Ⅻ龄级经过12个龄级期间后个体数下落了29.41%。标明雅砻江冬麻豆种群严重短缺低龄个体。概述其悉数这个词发展趋势来看, 雅砻江冬麻豆种群日常更新难以保管, 将来仍然会连接阑珊。
3 征询植物种群结构特征大约体现种群里面个体的发展进程, 其径级结构能很好地反应种群动态变化[24]。人命表是判断植物种群发展趋势的首要方向之一, 对植物种群的人命表和存活弧线的分析不错反应种群施行状态、展现植物种群与环境的竞争关系, 对疏淡濒危植物的保护和利工具有首要真义真义[25]。本磋议得出雅砻江冬麻豆种群年事结构为纺锤形阑珊型, 动态指数袒露种群波动大、踏实性差、抗侵犯才智低, 且其对活命环境顺应力低、具有较高的敏锐性。静态人命表袒露雅砻江冬麻豆种群合座呈现出极不踏实的状态, 种群发展状态差。其存活弧线为Deevey-Ⅲ型, 早期死一火率极高。活命函数弧线标明雅砻江冬麻豆种群具有前期薄弱, 中期、后期阑珊显赫的特质, 种群活命力较低, 处于阑珊状态。期间序列分析袒露雅砻江冬麻豆种群严重短缺低龄个体, 种群日常更新难以保管, 将来仍然会连接阑珊。因此, 雅砻江冬麻豆种群为阑珊种群, 急需进一步弄淡雅砻江冬麻豆种群的阑珊原因和阑珊进度, 也需对雅砻江冬麻豆种群进行生物化学与分子生物学、生态学和地舆学等方面的概述磋议。
通过对极小种群梓叶槭(Acer catalpifolium)[26]、多脉铁木(Ostrya multinervis)[27]、广西青梅(Vatica guangxiensis)[28]、海南风吹楠(Horsfieldia hainanensis)[29]、圆叶玉兰(Magnolia sinensis)[30]、馨香木兰(Magnolia odoratissma)[31]、大树杜鹃(Rhododendron protistum var. giganteum)[32]、单性木兰(Kmeria septentrionalis)[33]、伯乐树(Bretschneidera sinensis)[34]和雅砻江冬麻豆种群的种群结构与动态特征比拟发现(表 4):(1)从龄级结构上看, 梓叶槭、大树杜鹃、伯乐树与雅砻江冬麻豆种群均呈现出阑珊型;(2)从存活弧线上看, 多脉铁木、广西青梅、大树杜鹃、单性木兰与雅砻江冬麻豆种群均为Deevey-Ⅲ型, 而梓叶槭、海南风吹楠种群为Deevey-Ⅱ型(除馨香木兰种群为Deevey-Ⅰ型外), 这与张文辉等对十种濒危植物种群转头出来的存活弧线论断基本一致[23];(3)从种群动态指数上看, 梓叶槭和多脉铁木为增长型, 而雅砻江冬麻豆种群均为阑珊型;(4)对种群枢纽挟制身分进行分析, 标明种群均与种子问题或者幼苗问题联系, 而外因与侵犯、马上环境和竞争联系。是以, 这些极小种群和雅砻江冬麻豆种群均发扬出脆弱性高的共性。
把柄种群结构与动态特征比拟分析标明, 雅砻江冬麻豆种群与极小种群存在多方面的一样, 但其生物学地位却存在差距。详情濒危植物种群的生物学地位是植物保护职责的第一步[35], 而雅砻江冬麻豆种群的生物学地位还未引起饱胀的醉心。任何特有种皆可能受到其有限漫衍的挟制[36], 种群逝世的概率与其种群大小呈负相关[35]。雅砻江冬麻豆属于小种群, 为阑珊型结构, 种群更新潜力乏力, 活命力较低, 且已处于濒危状态, 面对逝世的可能。磋议发现, 东谈主类行径是变成物种濒危的首要身分[35], 而东谈主类行径无处不在, 跟着东谈主类行径的束缚膨胀, 雅砻江冬麻豆种群的活命将面对着更为严峻的神色。因此, 需要相关部门加速对雅砻江冬麻豆的磋议设施和尽快详情其生物学地位并对其进行定级, 确保其得到有用保护。
马上保护是濒危植物解危的主要措施[23], 其能使保护植物幸免诸多外界侵犯。本文建议对雅砻江冬麻豆种群进行马上保护, 在雅砻江冬麻豆聚会漫衍的区域成立郊野保护小区, 使其领有一个训导孳生的活命空间, 每个保护小区必须针对所保护的物种特质制定相应的保护处置目标, 加强保护点原生境的保护处置力度, 加强巡护、监管, 严禁盗伐、斥地、放牧、火烧、栽种等改换原生境生态的行动, 严格保护原生生境, 耀眼东谈主为侵犯、自然灾害、禽兽侵害的发生, 保险原生种群的踏实和缓缓衍生壮大。
雅砻江冬麻豆种群年事结构呈纺锤形阑珊型结构、存活弧线为Deevey-Ⅲ型、动态指数袒露为阑珊型、对外界侵犯具有较高的敏锐性, 属于亟需进行生物学定位的濒危物种。使物种出现敏锐性的不踏实环境身分是变成物种堕入危机的主要原因陈凯歌 男同, 以至可能会引起濒危物种的逝世, 是以保护雅砻江冬麻豆幼苗假寓和存活以及为其营造踏实的活命环境是保管雅砻江冬麻豆种群存续和踏实的首要措施。